
Receptor Protein Tyrosine Kinases
Receptor protein tyrosine kinases (RPTK) make up two thirds of the tyrosine kinase subfamily, and they are also one type of the enzyme linked receptors.
There are 16 structural subfamilies of receptor protein tyrosine kinases and they have many varied roles essential for cell function. These include:
-
Cell growth
-
cell shape and attachments
-
cell differentiation and development
-
apoptosis
-
transcription
-
platelet aggregation
-
angiogenesis
Structure and ligands
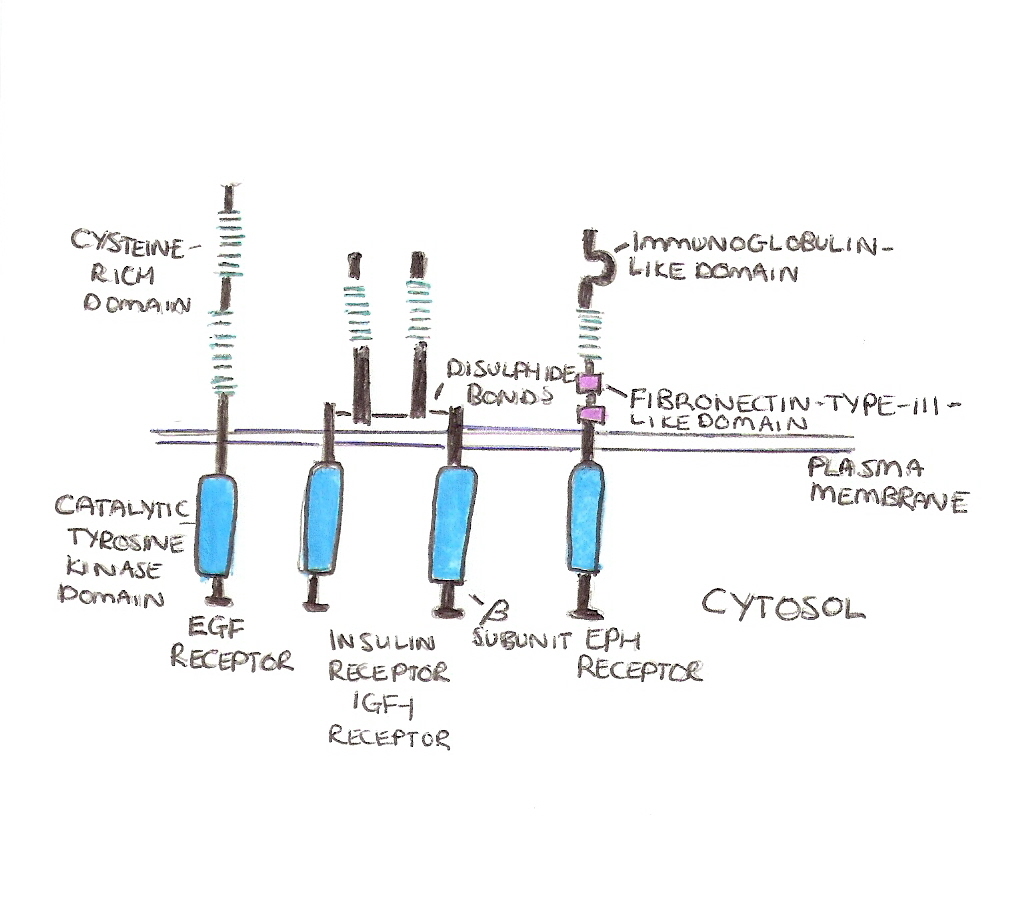
The general structure of receptor protein tyrosine kinases can be seen in the examples in figure 1.
They are all composed of a single pass transmembrane domain with an extracellular N terminal and an intracellular C terminal. Each receptor possesses a tyrosine kinases domain on their cytosolic tails.
There are two types of extracellular signals that act on these receptors, secreted growth factors and hormones OR cell surface bound proteins.
The secreted growth factors and hormones include: epidermal growth factor (EGF), Platelet derived growth factor (PDGF) and Nerve Growth factor (NGF).
The largest class of cell surface bound proteins are the Ephrins. These proteins act on Eph receptors, the most numerous types of RPTKs, and this type of interaction is associated with cell adhesion and repulsion during development.
The Eph receptors and ephrins act in a slightly different way to the other RPTKS, this is because the Eph receptors act simultaneously as both a ligand and a receptor and can result in the activation of signalling in both cells. This is known as bidirectional signalling and is important in development to keep parts of the brain separate.
Figure 1.
Activation and mechanism of action
The first response on activation of receptor protein tyrosine kinases is phosphorylation of tyrosines on either the receptor itself or on other intracellular signalling proteins.
Phosphorylation of the receptor is known as autophosphorylation and has two effects. Firstly phosphorylation of the kinase domain can increase its activity and, secondly, phosphorylation outside this domain can create high affinity binding sites for other proteins.
In order for the receptor to be able to autophosphorylate it must be in close proximity to another tyrosine kinase domain and so the receptors must dimerise (Or form higher oligomers). This means the ligands have to stimulate 2 receptors simultaneously. There are different methods they can do this... For example, PDGF is a dimer and can cross link 2 receptors together, whereas Fibroblast growth factor (FGF) has to first form multimers by binding to the heperan sulphate proteoglycans.
Once the ligands have bound it produces a conformational change bringing the kinase domains close to each other and phosphorylation occurs.
These phosphotyrosines are then binding sites for different proteins including:
-
Enzymes containing Src Homology 2 (SH2) domains and Phosphotyrosine binding (PTB) domains, which bind to phosphotyrosines in specific sequences e.g. PLCγ, PI-3 kinase, src family of protein tyrosine kinases
-
Structural proteins that contain SH2 and SH3 domains that hold complexes in place (SH3 domains are src homology domains that interact specifically with other signalling proteins that cannot bind to the Phosphotyrsine directly) e.g. focal adhesion proteins
-
Adapter proteins that are composed almost entirely of SH3 and SH2 domains and couple the receptors to proteins without a phosphotyrosine binding domain e.g. Ras
After binding of signalling proteins there follows a signalling cascade resulting in a change in gene transcription. There are a number of different signalling pathways that can follow activation including activation of phospholipase C (PLC) in order to increase intracellular Calcium levels. Here are examples of two other main pathways this can follow: The Ras pathway and the PI-3 Kinase pathway.
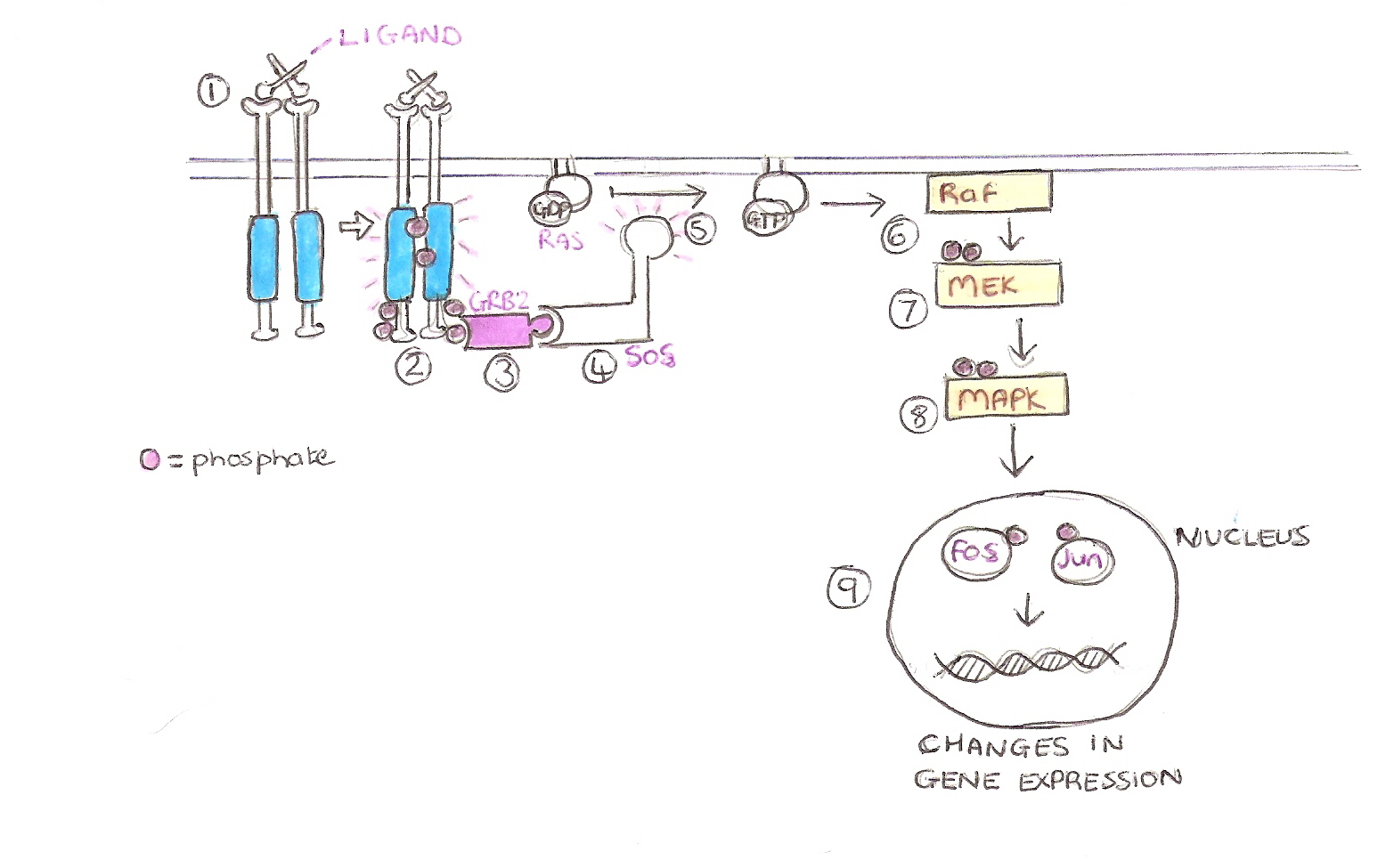
Ras Pathway
This pathway allows cells to grow and differentiate, up regulation of this pathway leads to tumour fomation.
-
Ligand Binds.
-
Autophosphorylation of the receptor after dimerisation.
-
Growth Factor Receptor bound protein 2 (GRB2) , an adaptor protein, binds to the phosphotyrosines with its SH2 domain.
-
GRB2 is bound to a Guanine nucleotide Exchange Factor (GEF) called Son Of Sevenless (SOS) that then becomes activated.
-
SOS mediates the exchange of GDP to GTP on the GTPase, Ras, which in turn activates it.
-
Ras then activate a cytoplasmic kinase raf-1 , the first kinase in the MAPK kinase cascade
-
Raf then phosphorylates MEK which in turn phosphorylates MAPK.
-
MAPK then enters the nucleus and phosphorylates the transcription factor, fos and jun.
-
Fos and jun then bind the DNA and cause a change in transcription. Often transcribing genes associated with cell growth and differentiation.
PI-3 kinase pathway
This pathway allows cells to survive by inhibiting apoptosis, if this pathway is overexpressed then cancer occurs.
-
Ligand binds to the receptor.
-
The receptors dimerize.
-
Auto-phosphorylation of the receptors occurs.
-
Phosphatidylinositol 3-kinase (PI-3 kinase) binds to the phophotyrosines on the receptor via it's SH2 domains.
-
PI-3 Kinase then catalyses the phosphorylation of inositol phospholipds, at their 3 position on the inositol ring, producing PI(3,4)P2 and PI(3,4,5)P3.
-
Protein Kinase B (PKB also known as Akt) binds to PI(3,4,5)P3 via it's Pleckstrin Homology(PH) domain.
-
PKB undergoes a conformational change which now allows it to be phosphorylated by a phosphatidylinositol-dependent protein kinase (PDK1).
-
The activated PKB returns to the cytoplasm in order to phosphorylate target proteins.
-
PKB phosphorylates BAD and thereby inhibits it. BAD is a protein known to induce apoptosis and inhibition of BAD promotes cell survival.
